




Arachnologiczny zespół badawczy
 dr Teresa Napiórkowska, prof. UMK; dr Julita Templin
dr Teresa Napiórkowska, prof. UMK; dr Julita Templin
Eksperymentalne badania z zakresu embriogenezy, postembriogenezy i teratogenezy
Morfometria układu nerwowego Tegenaria atrica
Regeneracja odnóży i struktur pochodzenia przysadkowego u Tegenaria atrica
Eksperymentalne badania z zakresu embriogenezy, postembriogenezy i teratogenezy
 Materiałem badawczym, na którym prowadzone są badania laboratoryjne jest pająk Tegenaria atrica C.L.Koch należący do rodziny Agelenidae (lejkowcowatych). Jest to jeden z gatunków licznie reprezentujących naszą krajową arachnofaunę, a zarazem jeden z najbardziej rozpoznawalnych gatunków ze względu na duże rozmiary ciała osobników dojrzałych płciowo i charakterystyczne ubarwienie. Jest to pająk typowo synantropijny, towarzyszący człowiekowi zwykle w miesiącach zimowych, kiedy to ze środowiska naturalnego migruje do piwnic, zabudowań gospodarczych czy mieszkań w poszukiwaniu korzystnych warunków do życia, a szczególnie dogodnych miejsc do rozrodu.
Materiałem badawczym, na którym prowadzone są badania laboratoryjne jest pająk Tegenaria atrica C.L.Koch należący do rodziny Agelenidae (lejkowcowatych). Jest to jeden z gatunków licznie reprezentujących naszą krajową arachnofaunę, a zarazem jeden z najbardziej rozpoznawalnych gatunków ze względu na duże rozmiary ciała osobników dojrzałych płciowo i charakterystyczne ubarwienie. Jest to pająk typowo synantropijny, towarzyszący człowiekowi zwykle w miesiącach zimowych, kiedy to ze środowiska naturalnego migruje do piwnic, zabudowań gospodarczych czy mieszkań w poszukiwaniu korzystnych warunków do życia, a szczególnie dogodnych miejsc do rozrodu.
 Aktywność rozrodcza bowiem przypada na okres jesienno ?zimowy, po czym z wiosną, wraz z podwyższeniem się temperatur dobowych opuszcza zajmowane do tej pory środowisko przenosząc się na otwartą przestrzeń, ale i wtedy przebywa w pobliżu zabudowań mieszkalnych. Stosunkowo szeroki zasięg występowania, łatwość pozyskania go ze środowiska naturalnego, szybka adaptacja do warunków laboratoryjnych, jak i niekłopotliwe prowadzenie hodowli nawet dla początkującego arachnologa, czyni go doskonałym obiektem dla prostych obserwacji behawioralnych, jak i bardziej zawansowanych projektów badawczych. Ponieważ samice składają jaja w kilku, a nawet kilkunastu złożach (maksymalnie 16), w których może znajdować się duża liczba sypkich, nieposklejanych jaj (do 150 w jednym, początkowym złożu), można je dowolnie sortować i przebierać w celu obserwacji wybranych, pojedynczych embrionów. Jest to bardzo istotna cecha, którą należy brać pod uwagę podczas planowania eksperymentu, gdzie istotna jest powtarzalność doświadczenia na embrionach tego samego osobnika.
Aktywność rozrodcza bowiem przypada na okres jesienno ?zimowy, po czym z wiosną, wraz z podwyższeniem się temperatur dobowych opuszcza zajmowane do tej pory środowisko przenosząc się na otwartą przestrzeń, ale i wtedy przebywa w pobliżu zabudowań mieszkalnych. Stosunkowo szeroki zasięg występowania, łatwość pozyskania go ze środowiska naturalnego, szybka adaptacja do warunków laboratoryjnych, jak i niekłopotliwe prowadzenie hodowli nawet dla początkującego arachnologa, czyni go doskonałym obiektem dla prostych obserwacji behawioralnych, jak i bardziej zawansowanych projektów badawczych. Ponieważ samice składają jaja w kilku, a nawet kilkunastu złożach (maksymalnie 16), w których może znajdować się duża liczba sypkich, nieposklejanych jaj (do 150 w jednym, początkowym złożu), można je dowolnie sortować i przebierać w celu obserwacji wybranych, pojedynczych embrionów. Jest to bardzo istotna cecha, którą należy brać pod uwagę podczas planowania eksperymentu, gdzie istotna jest powtarzalność doświadczenia na embrionach tego samego osobnika.
Eksperymentalne badania z zakresu embriogenezy i postembriogenezy pająka Tegenaria atrica.
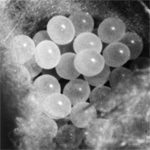 Na sukcesywnie uzyskiwanych embrionach w sezonach rozrodczych badany był całkowity czas embriogenezy oraz normalny przebieg i okres trwania następujących po sobie, kolejnych stadiów rozwojowych. Obserwacje prowadziliśmy posługując się lupą i używając oleju parafinowego, w którym zanurzaliśmy losowo wybrane embriony. Olej doskonale prześwietlał osłony jajowe i co było szczególnie istotne, nie hamował procesów rozwojowych. Ustaliliśmy, że najbardziej optymalne warunki dla rozwoju jaj to temperatura 21-230C i wilgotność względna 70%. W takich warunkach embriogeneza trwa 20 dni. Obserwowaliśmy wszystkie stadia rozwojowe rozłożone w czasie, a więc od stadium rozet, poprzez wczesną i późną blastodermę, dysk zarodkowy, płytkę pierwotną i bruzdę pierwotną, cumulus, pole dorsalne, zakładanie się odnóży, blastokinezę aż do wylęgu. Ustaliliśmy, że embriogeneza u tego pająka nie odbiega zasadniczo od modelu opisanego u innych gatunków pająków, np. Torania variata czy Cupiennius salei. Badaliśmy również czas trwania postembriogenezy. Okazało się, że zależy on głównie od temperatury, wilgotności otoczenia oraz ilości i jakości pożywienia. Stwierdziliśmy, że kolejne stadia rozwojowe charakteryzują się różną długością trwania. Im starsze stadium nimfalne, tym dłuższy czas trwania. Ustaliliśmy, że w warunkach laboratoryjnych osiągnięcie dojrzałości płciowej następowało po 10-12 miesiącach.
Na sukcesywnie uzyskiwanych embrionach w sezonach rozrodczych badany był całkowity czas embriogenezy oraz normalny przebieg i okres trwania następujących po sobie, kolejnych stadiów rozwojowych. Obserwacje prowadziliśmy posługując się lupą i używając oleju parafinowego, w którym zanurzaliśmy losowo wybrane embriony. Olej doskonale prześwietlał osłony jajowe i co było szczególnie istotne, nie hamował procesów rozwojowych. Ustaliliśmy, że najbardziej optymalne warunki dla rozwoju jaj to temperatura 21-230C i wilgotność względna 70%. W takich warunkach embriogeneza trwa 20 dni. Obserwowaliśmy wszystkie stadia rozwojowe rozłożone w czasie, a więc od stadium rozet, poprzez wczesną i późną blastodermę, dysk zarodkowy, płytkę pierwotną i bruzdę pierwotną, cumulus, pole dorsalne, zakładanie się odnóży, blastokinezę aż do wylęgu. Ustaliliśmy, że embriogeneza u tego pająka nie odbiega zasadniczo od modelu opisanego u innych gatunków pająków, np. Torania variata czy Cupiennius salei. Badaliśmy również czas trwania postembriogenezy. Okazało się, że zależy on głównie od temperatury, wilgotności otoczenia oraz ilości i jakości pożywienia. Stwierdziliśmy, że kolejne stadia rozwojowe charakteryzują się różną długością trwania. Im starsze stadium nimfalne, tym dłuższy czas trwania. Ustaliliśmy, że w warunkach laboratoryjnych osiągnięcie dojrzałości płciowej następowało po 10-12 miesiącach.
Poznanie wpływu zmian temperatury na rozwój embrionalny pająka Tegenaria atrica oraz badanie mechanizmów teratogennego działania tego czynnika
 W naszym laboratorium prowadzimy badania z zakresu teratologii eksperymentalnej. Okazuje się bowiem, że nie tylko czynniki fizyczne, fizykochemiczne i czysto chemiczne doprowadzają do bardzo głębokich deformacji ciała, ale również temperatury niższe i wyższe od optimum dla rozwoju zarodkowego powodują uszkodzenia embrionów. Zarodki pająków uzyskane w naszym laboratorium poddajemy naprzemiennemu działaniu dwóch temperatur: 14 i 320C przez okres ok. 10 dni, tj. do momentu pojawiania się na smudze zarodkowej pierwszych metamerów ciała. W ten sposób potraktowane embriony nie rozwijają się prawidłowo i w rezultacie obserwujemy różnorodne anomalie w obrębie prosomy i opistosomy.
W naszym laboratorium prowadzimy badania z zakresu teratologii eksperymentalnej. Okazuje się bowiem, że nie tylko czynniki fizyczne, fizykochemiczne i czysto chemiczne doprowadzają do bardzo głębokich deformacji ciała, ale również temperatury niższe i wyższe od optimum dla rozwoju zarodkowego powodują uszkodzenia embrionów. Zarodki pająków uzyskane w naszym laboratorium poddajemy naprzemiennemu działaniu dwóch temperatur: 14 i 320C przez okres ok. 10 dni, tj. do momentu pojawiania się na smudze zarodkowej pierwszych metamerów ciała. W ten sposób potraktowane embriony nie rozwijają się prawidłowo i w rezultacie obserwujemy różnorodne anomalie w obrębie prosomy i opistosomy.
 Należą do nich takie anomalie jak np.: oligomelia (brak odnóża/odnóży), polimelia (obecność dodatkowego odnóża/odnóży), heterosymelia (zrost odnóży leżących obok siebie), symelia (zrost odnóży należących do tej samej pary), schistomelia (rozwidlenie odnóży) czy dwugłowość. Niektóre z nich, np. obecność przysadki na petiolus lub w miejscu płucotchawki mają charakter atawizmów. Jest to temat, który prowadzony jest od wielu lat, a mimo to jest jeszcze wiele do zrobienia. Wciąż poszukujemy mechanizmów powstawania deformacji rozwojowych na różnych poziomach organizacji ciała.
Należą do nich takie anomalie jak np.: oligomelia (brak odnóża/odnóży), polimelia (obecność dodatkowego odnóża/odnóży), heterosymelia (zrost odnóży leżących obok siebie), symelia (zrost odnóży należących do tej samej pary), schistomelia (rozwidlenie odnóży) czy dwugłowość. Niektóre z nich, np. obecność przysadki na petiolus lub w miejscu płucotchawki mają charakter atawizmów. Jest to temat, który prowadzony jest od wielu lat, a mimo to jest jeszcze wiele do zrobienia. Wciąż poszukujemy mechanizmów powstawania deformacji rozwojowych na różnych poziomach organizacji ciała.
Rozwój aparatu jadowego u pająka Tegenaria atrica C.L.Koch w postembriogenezie.
Na aparat jadowy pająków składa się narząd raniący w postaci szczękoczułków oraz gruczoły jadowe. Celem podjęcia tych badań było oszacowanie zmian, jakim podlega ten aparat w czasie całego rozwoju począwszy od stadium larwalnego a na osobnikach dojrzałych kończąc. Złożyliśmy, że jego rozwój będzie skorelowany z powiększaniem się całego głowotułowia co potwierdziły szczegółowe badania oparte na analizie histometrycznej preparatów, które wykonaliśmy z osobników będących w początkowych stadiach rozwoju postembrionalnego. Można więc stwierdzić, że sukcesywny rozwój tego aparatu jest związany z coraz to większym zapotrzebowaniem na bardziej zróżnicowany pokarm.
Badania biometryczne okryw kutikularnych Tegenaria atrica C.L.Koch w postembriogenezie
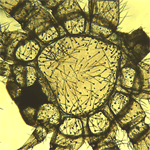 W eksperymentach wykorzystaliśmy zarówno osobniki prawidłowo wykształcone jak i teratologicznie zmienione, z oligomelią. Do badań wybraliśmy osobniki z brakiem jednej nogi chodowej, zawsze po tej samej stronie ciała. Wykorzystanie w doświadczeniu tylko jednego, określonego typu oligomelii (4/3, brak prawej nogi chodowej), zapewniło poprawne wyciąganie wniosków i uniknięcie błędów spowodowanych różnym tempem przebiegu procesów wzrostowych po obu stronach ciała pająka. Pomiary ciała pająków wykonaliśmy na podstawie sukcesywnie zbieranych egzuwiów, z których preparowaliśmy karapaksy i mostki. Ich pomiary (długość, szerokość, powierzchnia) wykonaliśmy za pomocą oprogramowania komputerowego Image J
W eksperymentach wykorzystaliśmy zarówno osobniki prawidłowo wykształcone jak i teratologicznie zmienione, z oligomelią. Do badań wybraliśmy osobniki z brakiem jednej nogi chodowej, zawsze po tej samej stronie ciała. Wykorzystanie w doświadczeniu tylko jednego, określonego typu oligomelii (4/3, brak prawej nogi chodowej), zapewniło poprawne wyciąganie wniosków i uniknięcie błędów spowodowanych różnym tempem przebiegu procesów wzrostowych po obu stronach ciała pająka. Pomiary ciała pająków wykonaliśmy na podstawie sukcesywnie zbieranych egzuwiów, z których preparowaliśmy karapaksy i mostki. Ich pomiary (długość, szerokość, powierzchnia) wykonaliśmy za pomocą oprogramowania komputerowego Image J  Na podstawie otrzymanych wyników, stwierdziliśmy, że karapaks i mostek osobników oligomelicznych jest zawsze mniejszy w porównaniu z kontrolą. Analiza statystyczna przy pomocy testu Studenta wykazała, że istniejące różnice w rozmiarach ciała pająków z grupy badanej i kontrolnej są istotne statystycznie. Jednakże osobniki oligomeliczne charakteryzują się bardzo silną tendencję zmierzającą do wyrównania zaistniałej anomalii. Przeprowadzone badania pokazały, że istnieją mechanizmy samoregulacji anomalii, które przejawiają się przyspieszeniem rozwoju postembrionalnego pająków oligomelicznych oraz kompensacja anomalii poprzez większe przyrosty ciała osobników z anomalią. Oczywistym wydaje się być fakt, iż oligomeliczne osobniki nie mają możliwości osiągnięcia rozmiarów ciała takich, jak pająki normalne. Oligomelia przejawia się nie tylko brakiem określonej liczby odnóży, ale również odpowiadającej im połówki metameru, a jak wiadomo, odtworzenie utraconego w czasie embriogenezy metameru ciała nie jest możliwe.
Na podstawie otrzymanych wyników, stwierdziliśmy, że karapaks i mostek osobników oligomelicznych jest zawsze mniejszy w porównaniu z kontrolą. Analiza statystyczna przy pomocy testu Studenta wykazała, że istniejące różnice w rozmiarach ciała pająków z grupy badanej i kontrolnej są istotne statystycznie. Jednakże osobniki oligomeliczne charakteryzują się bardzo silną tendencję zmierzającą do wyrównania zaistniałej anomalii. Przeprowadzone badania pokazały, że istnieją mechanizmy samoregulacji anomalii, które przejawiają się przyspieszeniem rozwoju postembrionalnego pająków oligomelicznych oraz kompensacja anomalii poprzez większe przyrosty ciała osobników z anomalią. Oczywistym wydaje się być fakt, iż oligomeliczne osobniki nie mają możliwości osiągnięcia rozmiarów ciała takich, jak pająki normalne. Oligomelia przejawia się nie tylko brakiem określonej liczby odnóży, ale również odpowiadającej im połówki metameru, a jak wiadomo, odtworzenie utraconego w czasie embriogenezy metameru ciała nie jest możliwe.
Morfometria układu nerwowego Tegenaria atrica
Struktura centralnego układu nerwowego w różnych przypadkach anomalii oraz morfometria tego układu u Tegenaria atrica w rozwoju postembrionalnym
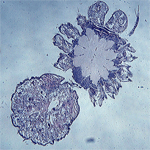 Analiza centralnego układu nerwowego oraz innych układów wewnętrznych oparta jest na preparatach histologicznych wykonanych metodą parafinową, po czym barwionych H+E. Dotyczy to szczególnie osobników z deformacjami typu oligomelii oraz polimelii. W różnorodnych przypadkach oligomelii odnóży (3/4, 3/3, 3/2 itd.) obserwowaliśmy brak odpowiednich zwojów nerwowych zgodnie z liczbą brakujących przysadek, natomiast w polimelii zawsze obecny był dodatkowy zwój lub kilka, odpowiednio do liczby wykształconych przysadek. Przeanalizowaliśmy również inne przypadki anomalii np. heterosymelię. W licznych przypadkach nie stwierdziliśmy zrostów ganglionów lecz było też kilka wyjątków, co zburzyło dotychczasowy pogląd o heterosymelii jako o zmianie niemetamerycznej. Interesujące są również wyniki dotyczące dwugłowości. W takich przypadkach stwierdziliśmy powiększanie się masy nerwowej o jeszcze jeden w pełni rozwinięty lub znacznie zdeformowany mózg. Podejmowane są również badania związane z morfometrią masy nerwowej u pająków ?zdrowych?, które obejmują cały okres postembrionalny. Interesuje nas zagadnienie dotyczące tempa rozwoju mózgu i masy podprzełykowej oraz ich składowych: korteksu i neuropilu w odniesieniu do tempa rozwoju prosomy. Badania opierają się na pełnych seriach skrawków histologicznych pająków będących w określonym stadium rozwojowym (larwa ? postać dorosła). Na podstawie obrazów cyfrowych, przy pomocy programu ImageJ obliczona zostaje powierzchnia interesujących nas składowych a następnie ich objętość. Otrzymane dotychczas wyniki wskazują, że u larw udział objętości całej masy nerwowej w objętości głowotułowia jest prawie równy 50%, w następnych stadiach natomiast następuje stopniowe obniżenie tego wskaźnika. U larw również, objętość mózgu przewyższa objętość masy podprzełykowej, a w trakcie dalszego rozwoju ma miejsce intensywne powiększanie się objętości masy podprzełykowej. Z porównania objętości korteksu i neuropilu podczas postembriogenezy wynika, że nieznacznie tylko wzrasta objętość korteksu, natomiast bardzo powiększa się objętość neuropilu. Wskazuje to na wytwarzanie się nowych połączeń dendrytycznych i powiększanie się istniejących już aksonów, przy stałej liczbie komórek tworzących korteks.
Analiza centralnego układu nerwowego oraz innych układów wewnętrznych oparta jest na preparatach histologicznych wykonanych metodą parafinową, po czym barwionych H+E. Dotyczy to szczególnie osobników z deformacjami typu oligomelii oraz polimelii. W różnorodnych przypadkach oligomelii odnóży (3/4, 3/3, 3/2 itd.) obserwowaliśmy brak odpowiednich zwojów nerwowych zgodnie z liczbą brakujących przysadek, natomiast w polimelii zawsze obecny był dodatkowy zwój lub kilka, odpowiednio do liczby wykształconych przysadek. Przeanalizowaliśmy również inne przypadki anomalii np. heterosymelię. W licznych przypadkach nie stwierdziliśmy zrostów ganglionów lecz było też kilka wyjątków, co zburzyło dotychczasowy pogląd o heterosymelii jako o zmianie niemetamerycznej. Interesujące są również wyniki dotyczące dwugłowości. W takich przypadkach stwierdziliśmy powiększanie się masy nerwowej o jeszcze jeden w pełni rozwinięty lub znacznie zdeformowany mózg. Podejmowane są również badania związane z morfometrią masy nerwowej u pająków ?zdrowych?, które obejmują cały okres postembrionalny. Interesuje nas zagadnienie dotyczące tempa rozwoju mózgu i masy podprzełykowej oraz ich składowych: korteksu i neuropilu w odniesieniu do tempa rozwoju prosomy. Badania opierają się na pełnych seriach skrawków histologicznych pająków będących w określonym stadium rozwojowym (larwa ? postać dorosła). Na podstawie obrazów cyfrowych, przy pomocy programu ImageJ obliczona zostaje powierzchnia interesujących nas składowych a następnie ich objętość. Otrzymane dotychczas wyniki wskazują, że u larw udział objętości całej masy nerwowej w objętości głowotułowia jest prawie równy 50%, w następnych stadiach natomiast następuje stopniowe obniżenie tego wskaźnika. U larw również, objętość mózgu przewyższa objętość masy podprzełykowej, a w trakcie dalszego rozwoju ma miejsce intensywne powiększanie się objętości masy podprzełykowej. Z porównania objętości korteksu i neuropilu podczas postembriogenezy wynika, że nieznacznie tylko wzrasta objętość korteksu, natomiast bardzo powiększa się objętość neuropilu. Wskazuje to na wytwarzanie się nowych połączeń dendrytycznych i powiększanie się istniejących już aksonów, przy stałej liczbie komórek tworzących korteks.
Regeneracja odnóży i struktur pochodzenia przysadkowego u Tegenaria atrica
 Tempo, jak i przebieg procesów regeneracyjnych śledziliśmy u pająków, u których uprzednio została wykonana bezkrwawa i bezpieczna dla życia pająków amputacja. Wykorzystaliśmy metodę przewiązek, które zakładane były osobnikom wybranych stadiów na określone wcześniej odnóże i miejsce na nim. Odrzucanie odpowiedniej części odnóża następowało podczas najbliższego linienia. Stwierdziliśmy, że przebieg regeneracji odpowiadał etapom ontogenezy odnóża chodowego. Kompletne odtworzenie odnóża następowało zwykle po 4 linieniach. Przeprowadzaliśmy również amputacje przysadek teratologicznie zmienionych. Wykazaliśmy, że epimorfoza jest zjawiskiem powszechnym i dotyczy w różnym stopniu zniekształconych przysadek. W przypadku polimelii oraz anomalii złożonych uruchomione zostają wyłącznie wstępne procesy regeneracyjne doprowadzające tylko do zagojenia się rany po tomii, natomiast odtworzenie amputowanej części odnóża następuje tylko w przypadku heterosymelii i schistomelii. Wyróżniliśmy trzy kolejne fazy epimorfozy, jak również obserwowaliśmy różne typy regeneratów w zależności od deformacji pierwotnej.
Tempo, jak i przebieg procesów regeneracyjnych śledziliśmy u pająków, u których uprzednio została wykonana bezkrwawa i bezpieczna dla życia pająków amputacja. Wykorzystaliśmy metodę przewiązek, które zakładane były osobnikom wybranych stadiów na określone wcześniej odnóże i miejsce na nim. Odrzucanie odpowiedniej części odnóża następowało podczas najbliższego linienia. Stwierdziliśmy, że przebieg regeneracji odpowiadał etapom ontogenezy odnóża chodowego. Kompletne odtworzenie odnóża następowało zwykle po 4 linieniach. Przeprowadzaliśmy również amputacje przysadek teratologicznie zmienionych. Wykazaliśmy, że epimorfoza jest zjawiskiem powszechnym i dotyczy w różnym stopniu zniekształconych przysadek. W przypadku polimelii oraz anomalii złożonych uruchomione zostają wyłącznie wstępne procesy regeneracyjne doprowadzające tylko do zagojenia się rany po tomii, natomiast odtworzenie amputowanej części odnóża następuje tylko w przypadku heterosymelii i schistomelii. Wyróżniliśmy trzy kolejne fazy epimorfozy, jak również obserwowaliśmy różne typy regeneratów w zależności od deformacji pierwotnej.
 Prowadziliśmy również eksperymenty, które polegały na amputacji kądziołków przędnych. W trakcie doświadczenia obserwowaliśmy blizny poamputacyjne, ewentualny proces regeneracyjny utraconych kądziołków przędnych, śmiertelność oraz możliwość wyrzucania przędzy u osobników poddanych tomii. U części osobników poddanych zabiegowi amputacji nie stwierdziliśmy regeneracji utraconych narządów, natomiast u pozostałych po kolejnych linieniach pojawiały się bardziej lub mniej zniekształcone kądziołki. Po zabiegu amputacji zewnętrznego aparatu przędnego pozostawała blizna, która była gładka lub pofałdowana. Stwierdziliśmy, że powierzchnia blizny nie ma wpływu na występowanie lub brak procesu regeneracyjnego. U części osobników mimo braku regeneracji kądziołków przędnych zaobserwowaliśmy bezładne wyrzucanie przędzy. Po wykonaniu preparatów glicerynowych stwierdziliśmy u takich osobników występowanie na polu poamputacyjnym pojedynczych struktur przędnych.
Prowadziliśmy również eksperymenty, które polegały na amputacji kądziołków przędnych. W trakcie doświadczenia obserwowaliśmy blizny poamputacyjne, ewentualny proces regeneracyjny utraconych kądziołków przędnych, śmiertelność oraz możliwość wyrzucania przędzy u osobników poddanych tomii. U części osobników poddanych zabiegowi amputacji nie stwierdziliśmy regeneracji utraconych narządów, natomiast u pozostałych po kolejnych linieniach pojawiały się bardziej lub mniej zniekształcone kądziołki. Po zabiegu amputacji zewnętrznego aparatu przędnego pozostawała blizna, która była gładka lub pofałdowana. Stwierdziliśmy, że powierzchnia blizny nie ma wpływu na występowanie lub brak procesu regeneracyjnego. U części osobników mimo braku regeneracji kądziołków przędnych zaobserwowaliśmy bezładne wyrzucanie przędzy. Po wykonaniu preparatów glicerynowych stwierdziliśmy u takich osobników występowanie na polu poamputacyjnym pojedynczych struktur przędnych.