![Katedra Ekologii i Biogeografii [fot. Katedra Ekologii i Biogeografii]](http://www.umk.pl/wiadomosci/serwisy_wp/gallery/1503/medium/KEiB.jpg "Katedra Ekologii i Biogeografii [fot. Katedra Ekologii i Biogeografii]")
Exciting theoretical results from our recent NCN grant:
Community assembly across temporal and spatial scales: unifying colonization and competition approaches.
The question how communities of animals and plant assembly has always been at the heart of ecological research. It was the seminal work of Jared Diamond who linked the different approaches to community assembly and provided a conception framework to study the pattern of species co-occurrences. Diamond?s vision of communities was based on the competitive exclusion principle and predicted mutual species exclusions of species with similar ecological requirements. This framework initiated a heavy and ongoing discussion about the importance of competition in ecological communities but also helped to develop appropriate methodological and statistical toolboxes to disentangle different processes of community assembly. Mainly in cooperation with Nick Gotelli from Vermont University we developed a comprehensive methodology for the study of community assembly based on a set of metrics and associated null models to study patterns of species co-occurrences and to link these patterns to environmental correlates and phylogenetic constraints. This toolbox is now widely used in community ecology.
Meta-analytical studies have shown that negative temporal and spatial species associations prevail within local communities, although by no means dominate. These findings indicate that in sufficiently stable environments niche and species interactions might drive community assembly in the tradition of Frederick Clements and that they might dominate over neutral (random) assembly as envisioned by Henry Gleason and formalized by Steven Hubbell.
Our results do not support this view.
Together with Franck Jabot from the Laboratoire d’Ingénierie pour les Syst?mes Complexes in Aubi?re, France and Nick Gotelli we were able to show that competition alone is not able to generate strong negative species associations within the observed range of local species richness. Instead we favour a largely neutral assembly.
But
 Diamond?s approach to species segregation was guided by the assumption of complete competitive exclusion as predicted by the strong version of Gause?s principle. However, incomplete exclusion and so-called competitive loops (A>B>C>A), also called competitive intransitivity, were shown to be common in multispecies communities Competitive intransitivity moderates the effects of competition, allowing weak competitors to coexist with strong ones. These approaches predict competitive intransitivity to generate stable species abundance distributions among sets of homogeneous sites but not species segregation as expected by competitive exclusion. Competitive intransitivity does not result in the prevalence of negative species associations. Therefore, major theories on community assembly predict contrasting patterns of species spatial or temporal distribution. Thus, competition might still be compatible with observed patterns of species associations.
Diamond?s approach to species segregation was guided by the assumption of complete competitive exclusion as predicted by the strong version of Gause?s principle. However, incomplete exclusion and so-called competitive loops (A>B>C>A), also called competitive intransitivity, were shown to be common in multispecies communities Competitive intransitivity moderates the effects of competition, allowing weak competitors to coexist with strong ones. These approaches predict competitive intransitivity to generate stable species abundance distributions among sets of homogeneous sites but not species segregation as expected by competitive exclusion. Competitive intransitivity does not result in the prevalence of negative species associations. Therefore, major theories on community assembly predict contrasting patterns of species spatial or temporal distribution. Thus, competition might still be compatible with observed patterns of species associations.
Our major aim was to develop a theoretical framework to predict community structure and to test respective hypotheses using empirical data sets at different spatial and temporal scales and to test this framework using high quality empirical data sets.
Our highlights from the recent grant
Task 1
 We succeeded in performing a detailed simulation study to infer patterns of species co-occurrences and abundances based on a neutral, ecological drift model on community assembly. This model was enriched by transitive and intransitive competitive interactions.
We succeeded in performing a detailed simulation study to infer patterns of species co-occurrences and abundances based on a neutral, ecological drift model on community assembly. This model was enriched by transitive and intransitive competitive interactions.
We asked
- How do changing competitive interactions influence patterns of species relative abundances?
- Is species segregation among patches linked to loops in competitive hierarchies?
- Does competitive looping according to the rock – paper – scissor game increase species diversity?
Highlighted by Oikos our results identified species dispersal (the movement among habitats) as the major trigger of community assembly. Competition is an auxiliary factor. Transitivity decreases, intransitivity increases local species richness.
A new technique to study species niches: environmental eigenvector ellipsoids

Ulrich W., Kryszewski W., Sewerniak P., Puchałka R., Strona G., Gotelli N.G. 2017. A comprehensive framework for the study of species co-occurrences, nestedness, and turnover. Oikos 126: 1607-1616.
Task 2
Astonishingly, despite of the long ecological tradition of studying competitive interactions and community assembly in natural systems, and despite of the enormous amount of theoretical approaches there is a clear lack of highly resolved field data to test existing hypotheses on plant and animal community structure.
We applied a new approach to plant data
High resolution false colour images allow for a precise geostatistical analysis of the spatial distribution of individual plants. This goes far beyond classical plant ecological and phytosociological analyses.
Papers are in preparation.
Task 3
Global databases contain many data and mysteries, but also many opportunities to resolve these mysteries around the biogeography of community assembly.
Pictures and paper
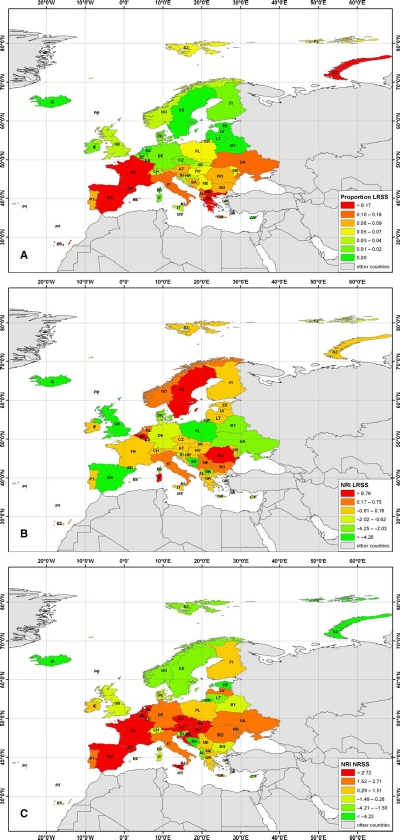
The phylogenetic community structure of European springtails
Fiera C., Habel J.C., Kunz M. Ulrich W. 2017. Large scale phylogenetic structure of European springtails (Collembola) depends on species range sizes and postglacial colonisation history. Biol. J. Linnean Soc. 120: 664-674
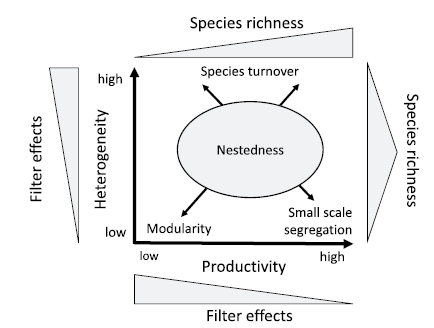
A general model of community assembly that emerges from our studies on understorey plant communities: